Cell:鄧宏魁與湯超等封面報(bào)道細(xì)胞重編程過程中的創(chuàng)新方法和理論
來(lái)源:生物360

近日,北京大學(xué)鄧宏魁研究組和湯超研究組合作,首次證明小鼠體細(xì)胞重編程可由調(diào)控分化的基因完成,并在此基礎(chǔ)上提出細(xì)胞命運(yùn)轉(zhuǎn)變的“蹺蹺板模型”。2013 年5月23日,該成果研究論文“Induction of pluripotency in mouse somatic cells with lineage specifiers”于《細(xì)胞》(Cell)期刊以封面文章形式在線發(fā)表。《細(xì)胞》同期還配發(fā)了希伯來(lái)大學(xué) Nissim Benvenisty 教授對(duì)于該工作的評(píng)論文章。
2006年,日本科學(xué)家 Shinya Yamanaka 發(fā)現(xiàn)向小鼠體細(xì)胞轉(zhuǎn)入胚胎干細(xì)胞特異因子(OCT4, SOX2, KLF4, c-MYC)可以完成體細(xì)胞的重編程。在此之后,細(xì)胞重編程領(lǐng)域普遍認(rèn)為向目標(biāo)細(xì)胞狀態(tài)的轉(zhuǎn)變需要靠在目標(biāo)狀態(tài)中特異高表達(dá)的因子的誘導(dǎo)。因此目前發(fā)現(xiàn)的 能夠在細(xì)胞重編程中發(fā)揮作用的因子均在胚胎干細(xì)胞中高表達(dá)并且與細(xì)胞干性維持緊密相關(guān)。
傳統(tǒng)觀點(diǎn)認(rèn)為分化因子與干性因子是相互拮抗,相互抑制的。干性基因在胚胎干細(xì)胞中高表達(dá),抑制分化基因;分化基因在胚胎干細(xì)胞中不表達(dá)或低表達(dá),高表達(dá)這些分化因子將抑制干性因子,破壞胚胎干細(xì)胞多能性的狀態(tài),導(dǎo)致其分化。
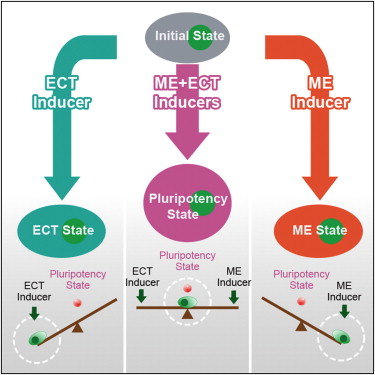
然而鄧宏魁研究組通過大規(guī)模篩選發(fā)現(xiàn),細(xì)胞重編程中至關(guān)重要的干性因子OCT4能夠被調(diào)控中內(nèi)胚層(ME)發(fā)育和分化的因子(如 GATA3,GATA6,PAX1)代替;SOX2能夠被調(diào)控外胚層(ECT)發(fā)育和分化的因子(如GMNN)代替。進(jìn)一步,湯超研究組根據(jù)這一發(fā)現(xiàn)創(chuàng)新 性地建立了“蹺蹺板模型”,來(lái)更好的理解中胚層基因和外胚層基因在重編過程中的相互抑制和相互平衡的關(guān)系,這種關(guān)系可能決定了細(xì)胞命運(yùn)的維持和改變。這個(gè) 模型提供了誘導(dǎo)體細(xì)胞重編程的其它方法的預(yù)測(cè),甚至還有一個(gè)出乎意料的模擬結(jié)果:如果同時(shí)過表達(dá)中內(nèi)胚層和外胚層基因,就可以達(dá)到平衡從而同時(shí)替代 SOX2和OCT4。而我們的實(shí)驗(yàn)結(jié)果也進(jìn)一步證實(shí)了這個(gè)可能,首次實(shí)現(xiàn)了用ME分化因子和ECT分化因子同時(shí)替代掉了細(xì)胞重編程過程中最關(guān)鍵的兩個(gè)干性 因子OCT4和SOX2。這一發(fā)現(xiàn)改變了向目標(biāo)細(xì)胞狀態(tài)的轉(zhuǎn)變需要用在目標(biāo)細(xì)胞狀態(tài)中高表達(dá)的因子的誘導(dǎo)的這一傳統(tǒng)觀點(diǎn),為研究細(xì)胞命運(yùn)轉(zhuǎn)變提供了新視 角,重新認(rèn)識(shí)了細(xì)胞重編程和細(xì)胞命運(yùn)決定的機(jī)制。
鄧宏魁研究組博士生舒健、吳晨、吳業(yè)濤與湯超研究組博士生李志遠(yuǎn)為共同第一作者,鄧宏魁研究組博士后趙揚(yáng)也在這一工作中發(fā)揮了關(guān)鍵作用。鄧宏魁、湯超均為北大-清華生命科學(xué)聯(lián)合中心成員。
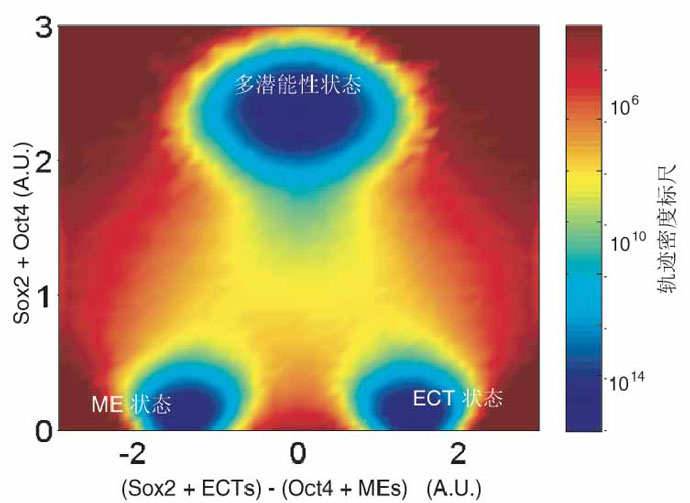
圖1.“Seesaw模型”的landscape圖
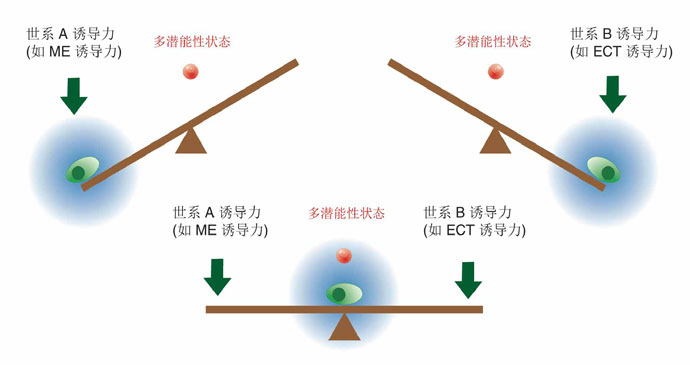
圖2. 細(xì)胞命運(yùn)決定的“蹺蹺板模型”
原文檢索:
Jian Shu, Chen Wu, Yetao Wu, Zhiyuan Li, Sida Shao, Wenhui Zhao, Xing Tang, Huan Yang, Lijun Shen, Xiaohan Zuo, Weifeng Yang, Yan Shi, Xiaochun Chi, Hongquan Zhang, Ge Gao, Youmin Shu, Kehu Yuan, Weiwu He, Chao Tang, Yang Zhao, Hongkui Deng. Induction of Pluripotency in Mouse Somatic Cells with Lineage Specifiers. Cell, 23 May 2013; DOI: 10.1016/j.cell.2013.05.001

